Scientifically accurate 3D model and animation of HIV

We created this 3D model and visualization of the HIV viral particle in 2009. In 2010, it was awarded first place and named Best Scientific Visualization at the Science and Engineering Visualization Challenge (SEVC), a competition organized by Science magazine and the National Science Foundation.
Since then, it has appeared on the covers of special issues of Nature Medicine and the International AIDS Vaccine Initiative (IAVI) Report, as well as in influential textbooks such as Vaccines and Principles of Virology. It has also been featured in The New York Times, Cell Picture Show by Cell Press, National Geographic, Wired, Popular Science, and other publications. The model received positive feedback from Nobel Prize winners, including Françoise Barré-Sinoussi.
We created this 3D model and visualization of the HIV viral particle in 2009. In 2010, it was awarded first place and named Best Scientific Visualization at the Science and Engineering Visualization Challenge (SEVC), a competition organized by Science magazine and the National Science Foundation.
Since then, it has appeared on the covers of special issues of Nature Medicine and the International AIDS Vaccine Initiative (IAVI) Report, as well as in influential textbooks such as Vaccines and Principles of Virology. It has also been featured in The New York Times, Cell Picture Show by Cell Press, National Geographic, Wired, Popular Science, and other publications. The model received positive feedback from Nobel Prize winners, including Françoise Barré-Sinoussi.
Project Goal
Create the most scientifically accurate and detailed 3D model of the HIV particle in atomic resolution.

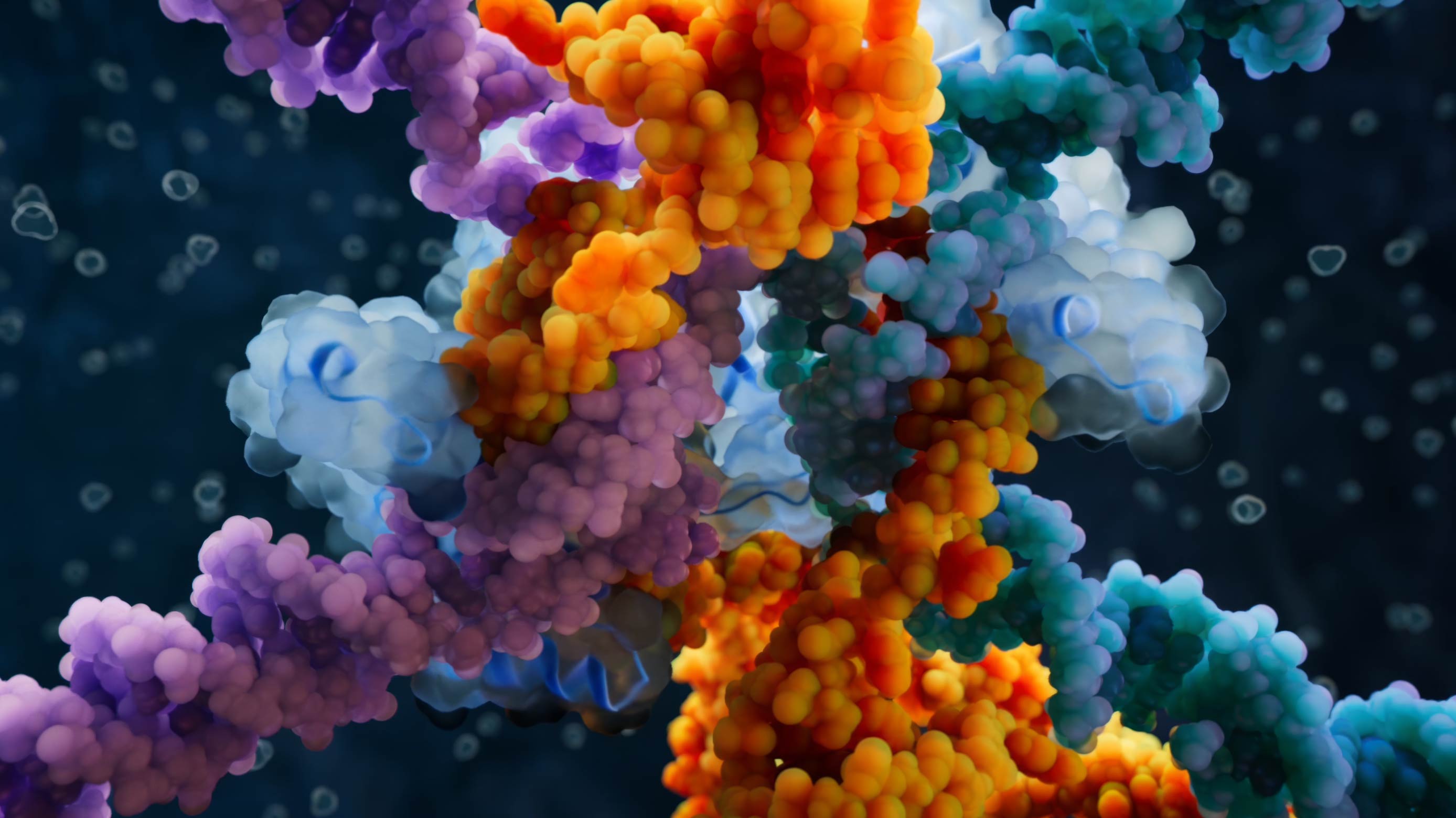
HIV whole model visualization

Surface proteins on the HIV particle
Envelope and capsid of the HIV particle
“Congratulations on your very successful 3D HIV model!”
“Congratulations on your very successful 3D HIV model!”

Nobel Prize in Medicine 2008, Pasteur Institute, Paris, France
Nobel Prize in Medicine 2008, Pasteur Institute, Paris, France
“Based on our experience, we highly recommend Visual Science as a reliable, trustworthy partner with excellent scientific, artistic and organizational expertise.”
“Based on our experience, we highly recommend Visual Science as a reliable, trustworthy partner with excellent scientific, artistic and organizational expertise.”

Executive Director Global HIV Vaccine Enterprise
Executive Director Global HIV Vaccine Enterprise

Nature Medicine special issue cover

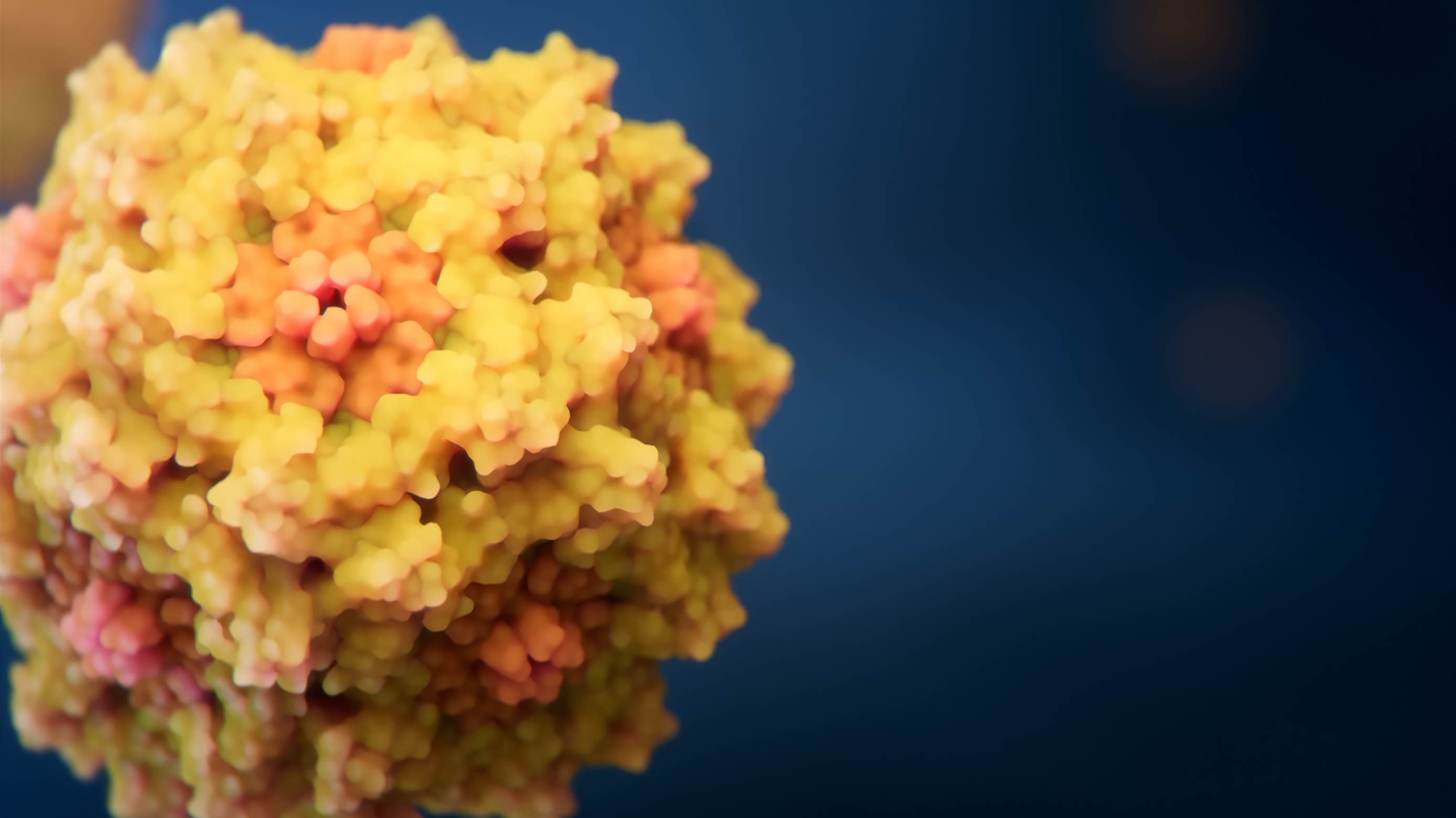
The particle core
Envelope proteins on the HIV surface
The Process
Animation type:
High-end 3D based on science grade molecular simulations
Project timline:
16 weeks
Find out which of our 7 scientific animation types and 20 subtypes work best for your project
This model summarizes results from more than 100 scientific publications published prior to 2009. Our team spent many weeks collecting and analyzing data, consulting with virologists, and assembling and refining the molecular structures of HIV components. None of the currently available scientific methods allow for obtaining an image of the entire virus particle at atomic resolution.
Nevertheless, hundreds of studies by researchers around the world have shed light on the structure and morphology of virion components and their interactions, making the creation of this model possible.
The first-ever molecular animation based on science-grade molecular simulations:
This complex and highly resource-intensive project was the first high-end molecular visualization we produced, and it set standards not only for our team but also for many of our colleagues in the industry. The depicted spatial configurations of 17 different viral and cellular proteins present in the HIV particle are strictly consistent with known 3D structures.
The viral membrane in the model includes approximately 160,000 lipid molecules of eight different types, in proportions consistent with those found in the HIV particle.
Full animation

Contact us
10/10
“
Fantastic team to partner with throughout the entire process. Excellent output and management.

Global Marketing Director, Pfizer
10/10
“
We are very pleased with Visual Science — they are a very responsive group to work with and the final product is exactly what we had envisioned.”

CEO, Atsena Therapeutics
10/10
“
The scientific expertise really showed in the discussions and final products. Everybody was responsive and great to work with, and the animations were both engaging and accurate!”

Program Manager, American Chemical Society
10/10
“
Visual Science team has just the right blend of scientific acumen and digital animation expertise to pull off these challenging projects.”

CEO, Apton Biosystems
9/10
“
Visual Science is a fantastic partner, capable of rendering the most complex science in compelling ways. They understood the science, and their production was excellent.”

VP, Head of Communications, Scorpion Tx
7/10
“
The structural biology models from Visual Science are stunning! Visual Science implemented our guidelines into beautiful scientific imagery for our website and scientific slide decks.”

Science Content Lead, Dewpoint Tx
9/10
“
I was very impressed with scientific knowledge of the Visual Science team, which led to a fantastic visualization of the MOA of our drug. Attention to scientific details was extraordinary.”

Vice President, R&D, Unicycive Tx
10/10
“
We were impressed by the scientific sophistication of the Visual Science team and their skill in capturing the concepts we wanted to communicate. The final video was exceptional.”

CSO, Enterin
10/10
“
The quality of the work from Visual Science was outstanding. They provide a bespoke service that was responsive to our requests and I would recommend them highly.”

CEO, UNITY Biotechnology
10/10
“
Beautiful work that is detailed and thoughtful. Great final output.”

Sr. Comms & IR Associate, Candel Tx
10/10
“
We were impressed by the Visual Science team’s ability to use exact protein structures to depict complex mechanisms in <90 seconds. The visuals are beautiful and tell a clear story.”

President & CEO, ROME Tx
10/10
“
Excellent scientific partner and first class imagery. This team runs a tight ship and the process is clear. The tech and final product are first rate.”

VP, Corp Dev, Revolution Medicines
10/10
“
Fantastic team to partner with throughout the entire process. Excellent output and management.
Global Marketing Director, Pfizer
10/10
“
We are very pleased with Visual Science — they are a very responsive group to work with and the final product is exactly what we had envisioned.”
CEO, Atsena Therapeutics
10/10
“
The scientific expertise really showed in the discussions and final products. Everybody was responsive and great to work with, and the animations were both engaging and accurate!”
Program Manager, American Chemical Society
10/10
“
Visual Science team has just the right blend of scientific acumen and digital animation expertise to pull off these challenging projects.”
CEO, Apton Biosystems
9/10
“
Visual Science is a fantastic partner, capable of rendering the most complex science in compelling ways. They understood the science, and their production was excellent.”
VP, Head of Communications, Scorpion Tx
7/10
“
The structural biology models from Visual Science are stunning! Visual Science implemented our guidelines into beautiful scientific imagery for our website and scientific slide decks.”
Science Content Lead, Dewpoint Tx
9/10
“
I was very impressed with scientific knowledge of the Visual Science team, which led to a fantastic visualization of the MOA of our drug. Attention to scientific details was extraordinary.”
Vice President, R&D, Unicycive Tx
10/10
“
We were impressed by the scientific sophistication of the Visual Science team and their skill in capturing the concepts we wanted to communicate. The final video was exceptional.”
CSO, Enterin
10/10
“
The quality of the work from Visual Science was outstanding. They provide a bespoke service that was responsive to our requests and I would recommend them highly.”
CEO, UNITY Biotechnology
10/10
“
Beautiful work that is detailed and thoughtful. Great final output.”
Sr. Comms & IR Associate, Candel Tx
10/10
“
We were impressed by the Visual Science team’s ability to use exact protein structures to depict complex mechanisms in <90 seconds. The visuals are beautiful and tell a clear story.”
President & CEO, ROME Tx
10/10
“
Excellent scientific partner and first class imagery. This team runs a tight ship and the process is clear. The tech and final product are first rate.”
VP, Corp Dev, Revolution Medicines
10/10
“
Fantastic team to partner with throughout the entire process. Excellent output and management.
Global Marketing Director, Pfizer
10/10
“
We are very pleased with Visual Science — they are a very responsive group to work with and the final product is exactly what we had envisioned.”
CEO, Atsena Therapeutics
10/10
“
The scientific expertise really showed in the discussions and final products. Everybody was responsive and great to work with, and the animations were both engaging and accurate!”
Program Manager, American Chemical Society
10/10
“
Visual Science team has just the right blend of scientific acumen and digital animation expertise to pull off these challenging projects.”
CEO, Apton Biosystems
9/10
“
Visual Science is a fantastic partner, capable of rendering the most complex science in compelling ways. They understood the science, and their production was excellent.”
VP, Head of Communications, Scorpion Tx
7/10
“
The structural biology models from Visual Science are stunning! Visual Science implemented our guidelines into beautiful scientific imagery for our website and scientific slide decks.”
Science Content Lead, Dewpoint Tx
9/10
“
I was very impressed with scientific knowledge of the Visual Science team, which led to a fantastic visualization of the MOA of our drug. Attention to scientific details was extraordinary.”
Vice President, R&D, Unicycive Tx
10/10
“
We were impressed by the scientific sophistication of the Visual Science team and their skill in capturing the concepts we wanted to communicate. The final video was exceptional.”
CSO, Enterin
10/10
“
The quality of the work from Visual Science was outstanding. They provide a bespoke service that was responsive to our requests and I would recommend them highly.”
CEO, UNITY Biotechnology
10/10
“
Beautiful work that is detailed and thoughtful. Great final output.”
Sr. Comms & IR Associate, Candel Tx
10/10
“
We were impressed by the Visual Science team’s ability to use exact protein structures to depict complex mechanisms in <90 seconds. The visuals are beautiful and tell a clear story.”
President & CEO, ROME Tx
10/10
“
Excellent scientific partner and first class imagery. This team runs a tight ship and the process is clear. The tech and final product are first rate.”
VP, Corp Dev, Revolution Medicines
10/10
“
Fantastic team to partner with throughout the entire process. Excellent output and management.
Global Marketing Director, Pfizer
10/10
“
We are very pleased with Visual Science — they are a very responsive group to work with and the final product is exactly what we had envisioned.”
CEO, Atsena Therapeutics
10/10
“
The scientific expertise really showed in the discussions and final products. Everybody was responsive and great to work with, and the animations were both engaging and accurate!”
Program Manager, American Chemical Society
10/10
“
Visual Science team has just the right blend of scientific acumen and digital animation expertise to pull off these challenging projects.”
CEO, Apton Biosystems
9/10
“
Visual Science is a fantastic partner, capable of rendering the most complex science in compelling ways. They understood the science, and their production was excellent.”
VP, Head of Communications, Scorpion Tx
7/10
“
The structural biology models from Visual Science are stunning! Visual Science implemented our guidelines into beautiful scientific imagery for our website and scientific slide decks.”
Science Content Lead, Dewpoint Tx
9/10
“
I was very impressed with scientific knowledge of the Visual Science team, which led to a fantastic visualization of the MOA of our drug. Attention to scientific details was extraordinary.”
Vice President, R&D, Unicycive Tx
10/10
“
We were impressed by the scientific sophistication of the Visual Science team and their skill in capturing the concepts we wanted to communicate. The final video was exceptional.”
CSO, Enterin
10/10
“
The quality of the work from Visual Science was outstanding. They provide a bespoke service that was responsive to our requests and I would recommend them highly.”
CEO, UNITY Biotechnology
10/10
“
Beautiful work that is detailed and thoughtful. Great final output.”
Sr. Comms & IR Associate, Candel Tx
10/10
“
We were impressed by the Visual Science team’s ability to use exact protein structures to depict complex mechanisms in <90 seconds. The visuals are beautiful and tell a clear story.”
President & CEO, ROME Tx
10/10
“
Excellent scientific partner and first class imagery. This team runs a tight ship and the process is clear. The tech and final product are first rate.”
VP, Corp Dev, Revolution Medicines

Contact us
Contact us
Visual Science is an award-winning medical animation and digital scientific communications company, trusted by leading biotech and pharmaceutical organizations since 2007, including J&J, Pfizer, Novartis, Roche, Takeda, Gilead, AbbVie, and 100+ others.
We specialize in science-grade MoA and MoD videos, medical animation, and scientific storytelling, as well as digital and AI-driven solutions for Medical Affairs, marketing, corporate communications, and investor relations.


© 2026, Visual Science. All rights reserved.



Visual Science is an award-winning medical animation and digital scientific communications company, trusted by leading biotech and pharmaceutical organizations since 2007, including J&J, Pfizer, Novartis, Roche, Takeda, Gilead, AbbVie, and 100+ others.
We specialize in science-grade MoA and MoD videos, medical animation, and scientific storytelling, as well as digital and AI-driven solutions for Medical Affairs, marketing, corporate communications, and investor relations.
© 2026, Visual Science. All rights reserved.
Visual Science is an award-winning medical animation and digital scientific communications company, trusted by leading biotech and pharmaceutical organizations since 2007, including J&J, Pfizer, Novartis, Roche, Takeda, Gilead, AbbVie, and 100+ others.
We specialize in science-grade MoA and MoD videos, medical animation, and scientific storytelling, as well as digital and AI-driven solutions for Medical Affairs, marketing, corporate communications, and investor relations.
© 2026, Visual Science. All rights reserved.